



藻類や植物がもつ「葉緑体」は細胞内共生によって、もともと葉緑体を持っていなかった生物が光合成をする生物を取り込み、やがては1つの生物になったことでもたらされた細胞内小器官です(図1)。
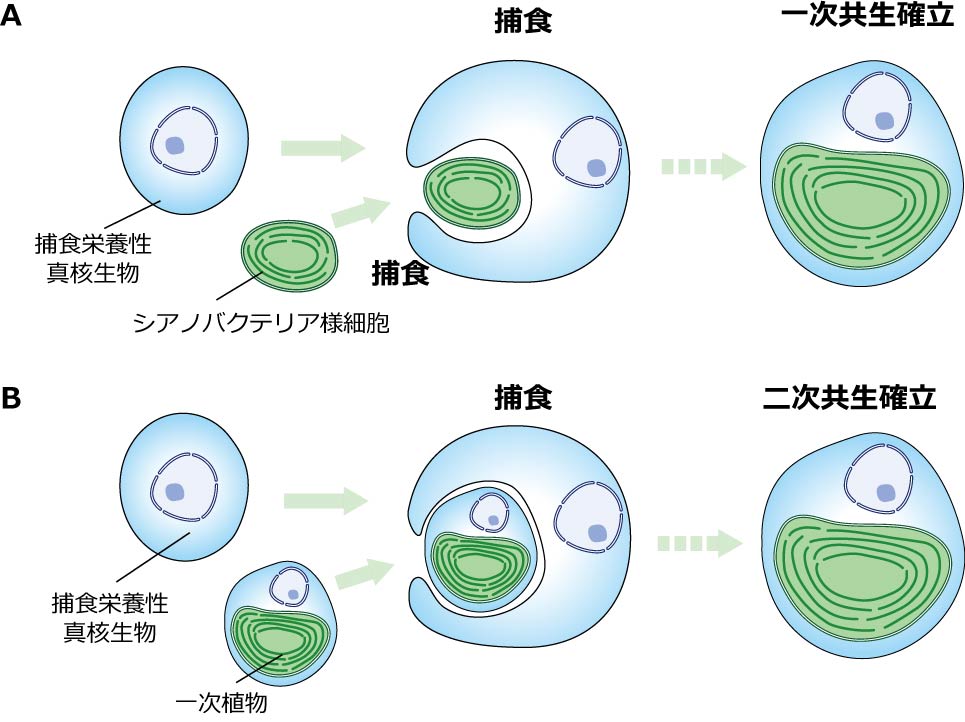
海洋・湖沼には、形態的・系統的に多様な「藻類」が生息していますが、それらはもともと他の生物を食べて生きていた生物が、細胞内共生によって葉緑体を獲得したことによって「藻類化」したことによって進化してきたと考えられています。しかしながら、そのダイナミックな進化がどのように起こってきたかという進化過程はほとんどわかっていません。
本研究室では、現在、他の藻類の葉緑体を盗む「盗葉緑体」現象を示す生物を主な研究対象とし、以下のテーマを掲げ、細胞内共生のメカニズム、藻類の進化を解明することを目指しています。
盗葉緑体現象のメカニズム解明
盗葉緑体現象とは、もともと葉緑体を持っていない生物が、葉緑体で光合成をして生きる生物を捕食し、その葉緑体を細胞内で一時的に盗んで使う現象を指します。私は研究開始当初から、盗葉緑体性の渦鞭毛藻Nusuttodinium(ヌスットディニウム)属を研究対象として使っています。属名のヌスットは日本語の盗人に由来します。この属の渦鞭毛藻類は、クリプト藻の葉緑体を盗んで使います。下の動画は渦鞭毛藻がクリプト藻を捕食する動画です。
ヌスットディニウムは、もともと葉緑体を持たない生物です。ヌスットディニウムは、上記のようにクリプト藻を捕食し、その葉緑体を細胞内で一時的に維持します。
ヌスットディニウムの仲間は数種類いることが知られていますが、海産のある種類は、葉緑体だけを細胞内で維持し、葉緑体の拡大は観察されません。クリプト藻の核は取り込まれたあとすぐに、消化されてしまいます(図2)。葉緑体もおよそ1、2週間で消化されてしまいます。
一方で淡水産のNusuttodinium aeruginosum(ヌスットディニウム・アエルギノーサム)は、クリプト藻の葉緑体の他に、クリプト藻の核も細胞内に維持し、なんと細胞全体にクリプト藻の葉緑体を拡大します。それだけにとどまらず、Nusuttodinium aeruginosumは細胞分裂時に盗んだ葉緑体を分裂させて、両方の娘細胞に受け継ぐことができます。この種は、クリプト藻1細胞さえあれば、100細胞以上に増えることができ、葉緑体を1ヶ月以上維持することができます。しかしながら、渦鞭毛藻が細胞分裂をする際には、クリプト藻の核は分裂せず、常に片方にのみ受け継がれます。細胞核を持っている細胞は分裂しても葉緑体のサイズを保っていますが、細胞核を失うと、葉緑体は分裂を繰り返すたびに小さくなっていってしまいます(図2)。このことから、葉緑体の拡大には、クリプト藻核が必要であることが示唆されていました。
この2種の比較から、細胞内に共生させた藻類を長く維持するためには、クリプト藻の核が必要であり、細胞内共生確立には、核を維持する進化段階が必要であるということが示唆されました。
最近になって、N. aeruginosumの盗んだ核には、遺伝子発現が保たれており、光化学系や炭酸固定に関連する遺伝子群が取り込まれる前のクリプト藻よりも発現上昇することが明らかとなっています。本研究室では、主にNusuttodiniumを用いて、このような一時的な共生関係を可能にするメカニズムを解明することを試みています。